舞茸中一种新型抗单纯疱疹病毒HSV-1蛋白的分离、
日期:2019-07-31 / 人气:
声明:本文摘自Antiviral Research 75 (2007) 250-257,中文名《抗病毒研究》,是一本关于病毒性疾病预防和治疗研究的杂志,由国际抗病毒研究协会(ISAR)官方出版,主要收录与人类及动物病毒感染控制相关的研究报告,综述以及评论等。为方便广大读者阅读,我们对论文摘要进行了中文翻译,译文仅供参考,如有不足之处,敬请指正。
【参考译文】
Isolation, identification and function of a novel anti-HSV-1 protein
from Grifola frondosa
舞茸中一种新型抗HSV-1蛋白的分离、鉴别及其功能确定
摘 要
用40%硫酸铵沉淀法和DEAE纤维素离子交换色谱法从舞茸子实体中提取一种新型的抗病毒蛋白,并命名为GFAHP。这种蛋白质能在体外抑制1型单纯疱疹病毒(HSV-1)的复制,其50%抑制浓度(IC50)为4.1μg/ml,治疗指数>29.3。在小鼠模型中,更高浓度的GFAHP(125和500μg/ml)也能显著减轻由HSV-1诱导的睑缘炎、新血管形成以及基质角膜炎。小鼠眼角膜外用GFAHP能显著减少病毒的产量(实验组为3.4log10PFU,对照组为4.19log10PFU)。我们证实,GFAHP在阻止HSV-1穿透进入肾细胞的同时能直接使HSV-1失活。凝胶电泳显示GFAHP的分子量为29.5kDa。用胰蛋白酶水解GFAHP后,从MALDI-TOF-MS测得的肽质量指纹谱(PMF)和电喷雾质谱法分析得知GFAHP的N端序列是一个含11个氨基酸的多肽,即NH2-REQDNAPCGLN-COOH,这与任何一种已知的氨基酸序列不匹配,这说明GFAHP很有可能是一种新型的抗病毒蛋白。据我们所知,这是关于舞茸的抗HSV蛋白的首个报道。
关键词:舞茸;蛋白质;抗病毒活性;鉴定;HSV-1
论文原文
Antiviral Research 75 (2007) 250–257
Isolation, identification and function of a novel anti-HSV-1 protein from Grifola frondosa
Chang-Qing Gu, Jun-Wen Li∗ , Fuhuan Chao, Min Jin, Xin-Wei Wang, Zhi-Qiang Shen
Institute of Environment and Health, 1 Dali Road, Tianjin 300050, PR China
Received 15 November 2006; accepted 22 March 2007
Abstract
A novel antiviral protein was purified from an extract of Grifola frondosa fruiting bodies using a procedure that included 40% ammonium sulfate precipitation and DEAE-cellulose ion exchange chromatography, and designated GFAHP. This protein inhibited herpes simplex virus type1 (HSV-1) replication in vitro with an IC50 value of 4.1μg/ml and a therapeutic index >29.3. Higher concentrations of GFAHP (125 and 500μg/ml) also significantly reduced the severity of HSV-1 induced blepharitis, neovascularization, and stromal keratitis in a murine model. Topical administration of GFAHP to the mouse cornea resulted in a significant decrease in virus production (mean virus yields: 3.4 log 10 PFU in the treated group and 4.19 log10 PFU in the control group). We proved that GFAHP directly inactivates HSV-1 while simultaneously inhibiting HSV-1 penetration into Vero cells. Gel electrophoresis showed that GFAHP had a molecular weight of 29.5kDa. GFAHP was tryptic digested and analyzed from the PMF of matrix assisted desorption ionization-time of flight mass spectrometry (MALDI-TOF-MS) and nanoelectrospray ionization tandem mass spectrometry. The N-terminal sequence of GFAHP consisted of an 11 amino acid peptide, NH2 -REQDNAPCGLN-COOH that did not match any known amino acid sequences, indicating that GFAHP is likely to be a novel antivirus protein. To our knowledge, this is the first report that characterizes an anti-HSV protein from G. frondosa.
© 2007 Elsevier B.V. All rights reserved.
Keywords: Grifola frondosa; Protein; Antiviral activity; Identification; HSV-1
1. Introduction
Fungi are an important source of anti-microbial and immuno-suppressive compounds. Many have also been tested for antibacterial activity and some mushrooms were recently shown to possess antiviral activity. The basidiomycete, Fomes fomentarius, for example, has antiviral activity against tobacco mosaic virus (Aoki et al., 1993) and an extract of the edible Japanese Lentinus edodes was found to inhibit replication of herpes simplex virus (HSV), western equine encephalitis virus, poliovirus, measles virus, and mumps virus (Sorimachi et al., 1990; Sarkar et al., 1993). The sterols from the edible mushroom, Hypsizigus marmoreus, have a potent inhibitory effect on Epstein-Barr virus activation induced by the tumor promoter 12-O-tetradecanoylphorbol-13-acetate (TPA) in Raji cells (Akihisa et al., 2005) and ganodermadiol and applanoxidic acid G from G.pfeifferi possess antiviral activity against influenza virus type A(IVA)(Mothana et al.,2003). In vitro antiviral activity against IVA was also shown for mycelial extracts of Kuehneromyces mutabilis (Mentel et al., 1994). Screening other mushroom species may help to identify additional antiviral factors.
The edible mushroom, G. frondosa, is a basidiomycete fungus belonging to the Polyporaceae family. Fruit bodies and liquid-cultured mycelium from this mushroom are reported to contain useful antitumor polysaccharides (Borchersetal.,1999; Adachi et al., 1994), and show some promise as immunomodulatory agents. They may also be beneficial for the treatment of hyperlipidemia, hypertension, and hepatitis (Mayell, 2001). It is reported that D-Fraction, a polysaccharide extracted from the maitake mushroom, G. frondosa, activates macrophages, dendritic cells, and T cells, inhibits tumor cell growth (Kodama et al., 2002, 2005; Inoue et al., 2002), enhances the cytotoxicity of NK cells by inducing the production of IL-12 (Kodama et al., 2002), and improves the symptoms and secondary diseases caused by HIV (Nanba et al., 2000). We showed that the D-Fraction also has the ability to inhibit HBV in vitro (Gu et al., 2006).
In general, it is the polysaccharides from mushrooms, like G. frondosa, that contain antiviral activity (Collins and Ng, 1997; Lee et al., 2003, 2004; Shi et al., 2006; Hsieh et al., 2006; Nie et al., 2006; Cui et al., 2007). In this study, we describe a novel protein from G. frondosa, named GFAHP which has antiviral activity. To our knowledge, this is the first isolation and characterization of a protein from this mushroom species that has antiviral function.
2.Materials and methods
2.1. Virus and cell culture
High titer virus stocks were grown in Vero cells as described previously (Grau et al., 1989). Dr. Wang at the National Institute for Viral Disease, Chinese Center for Disease Control and Prevention provided the HSV-1 strain, KOS, and the Vero cells. Vero cells were cultured in Dulbecco’s Modified Eagles Medium (DMEM) with 8% fetal bovine serum (FBS) at 37 ℃ with 5% CO2 . Virus stocks were prepared in cells fed with 2% FBS.
2.2. Isolation and identification of GFAHP
2.2.1. Preparation of G. frondosa extracts
Six hundred grams of G. frondosa mushrooms (Huayi Biotech Co., Henan, China) were homogenized with an equal amount of distilled water at room temperature (RT). The homogenate was heated at 60 ℃ for 12 h and centrifuged at 2500×g for 20min. The supernatent, designated as fraction 1, was lyophilized and resuspended in water to a concentration of 20mg/ml. Ammonium sulfate was slowly added to make a final concentration of 40%, stirred at 4 ℃ for 2 h, and centrifuged at 2000×g for 15 min. The precipitate was resuspended in deionized water, and both the precipitate (fraction 2) and the supernatent (fraction 3) were dialyzed against water for 2 days (four changes of water), lyophilized, and stored at −70 ℃. The dialysate was designated as fraction 4. As shown in Table 1, the antiviral activity was predominantly in fraction 2.

2.2.2. Isolation of GFAHP
A 37cm×1.3cm column was packed with DEAE-cellulose (Pharmacia Biotech) in distilled water, and an 8ml sample of fraction 2 (3.5mg/ml) in distilled water was passed through the column at a rate of 8ml/h and eluted with a linear NaCl gradient at 4 ℃. After removal of unabsorbed material (D1, designated fraction 5), the substance (D2, designated fraction 6) was eluted with 0.5M NaCl. Further substances were eluted with 1.0M NaCl (designated fraction 7) and 1.5 M NaCl (designated fraction 8).The absorbance of each fraction was measured at 280nm, and each was collected, dialyzed, and tested for anti-HSV-1 activity using the plaque reduction assay. The antiviral activity in fraction 6 was designated as G. frondosa antiviral protein, GFAHP.
2.2.3. Polyacrylamide gel electrophoresis
GFAHP was electrophoresed on a 15% denaturing-reducing sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE) as described previously (Visalli and Brandt, 1993). The molecular weight was determined by comparing electrophoretic mobility with molecular weight marker proteins purchased from Takara Biotechnology Company (Dalian, China).
2.2.4. Identification of GFAHP by mass spectrometry
Acetone precipitated GFAHP was electrophoresed and transferred to a PVDF membrane. The 29 kDa band containing the active material was cut from the membrane and sequenced from the amino terminus using the trypsin digestion procedure (Wang et al., 2006).
All mass spectra of MALDI-TOF-MS were obtained on a Bruker REFLEX III MALDI-TOF-MS (Bruker-Franzen, Bremen, Germany) in positive ion mode at an accelerating voltage of 20kV with a α-cyano-4-hydroxy cinnamic acid matrix. The spectra were internally calibrated using trypsin autolysis products. The peptide mass fingerprint (PMF) obtained was used
to search the SWISS-PROT and NCBInr database using the Mascot search engine with a tolerance of ±0.1 Da and one missed cleavage site.
ESI-MS/MS experiments were performed on a Q-TOF2 hybrid quadrupole/TOF mass spectrometer (Micromass, UK) with a nano flow Z-spray source. Peptide sequencing was performed using a palladium-coated borosilicate electrospray needle (Protana, Denmark). The mass spectrometer was used in positive ion mode with a source temperature of 80 ℃, and a potential of 800–1000V was applied to the Nanospray probe. The amino acid sequences of the peptides were deduced using the peptide sequencing program, MasSeq. The database search done by the Mascot search engine was completed using the data processed through MaxEnt3 and MasSeq. Each palladium-coated borosilicate electrospray needle (Protana, Odense, Denmark) was only used once.
2.3. GFAHP antiviral activity in vitro
2.3.1. Therapeutic effect of GFAHP on HSV-1 infection
To measure GFAHP activity, Vero cells were inoculated into wells of a 24-well plate at 2×105 cells/well and incubated overnight. Once confluent, the cells were infected with HSV-1 at a multiplicity of infection (MOI) of 2PFU/cell. Following a 1h adsorption period, the cells were rinsed twice with PBS (pH 7.4) and mixed with media containing different concentrations of GFAHP. Virus infected GFAHP treated, uninfected untreated, and uninfected GFAHP treated cells were tested in triplicate. At 48h post-infection (p.i.) the amount of infectious virus was measured by plaque assay and quantitated (Pirano and Brandt, 1999).
2.3.2. GFAHP pretreatment and direct virucidal activity
To determine the preventive effect of GFAHP on virus infected Vero cells, monolayers were incubated with 30 or 60μg/ml GFAHP for 24h at 37 ℃. After incubation, the cells were washed three times with PBS to remove unused GFAHP and infected with HSV-1 at an MOI of 2 to allow plaque formation. Untreated monolayers were used as controls (Chiang et al., 2002).
To investigate the direct virucidal effect of GFAHP on HSV-1, 1.5×106 PFU virus (0.1ml volume) was mixed with 0.1ml 60μg/ml GFAHP or 0.1ml medium and incubated at 37 ℃ for 1h. The mixture was diluted 1000× and assayed for residual virus by adsorbing for 1 h at 37 ℃. Inocula were removed and Vero cell cultures were washed twice prior to the addition of overlay medium.
2.3.3. HSV-1 cell attachment and penetration
Vero monolayers grown in 24-well plates were pre-chilled at 4℃ for 15min and infected with HSV-1 diluted in serum-free MEM to 200 PFU/ml for 1.0 h at 4 ℃ in the presence of GFAHP with 30 and 60 μg/ml or absence. Unadsorbed virus was then removed and plaques were allowed to develop at 37 ℃ for 72h (De Logu et al., 2000).
The penetration assay was performed as previously described with some modifications (Rosenthal and Peres, 1985; De Logu et al., 2000). HSV-1 stock was added to Vero cells in a 24-well culture plate and incubated at 4 ℃ for 2 h, and the temperature was abruptly increased to 37 ℃ to maximize penetration of the virus. Penetration proceeded for 1 h in the presence and absence of GFAHP. Monolayers were then treated with PBS, pH 3 for 1 min to neutralize any remaining attached virus and after several washes with serum-free medium, overlay media was added to promote plaque formation.
2.4. GFAHP antiviral activity in vivo
2.4.1. Experimental design of HSV-1 infection in mice and disease scoring
Five-week-old female BALB/c mice (Center of Animals, Academy of Military Medical Sciences, Beijing, China) were used for all studies. Mice were anesthetized by halothane inhalation. While under anesthesia, the right cornea was scratched three times vertically and four times horizontally with a sterile 27-gauge needle. A 10μl drop of DMEM containing 106 PFU of HSV-1 KOS was placed on the damaged cornea for 30 s. Excess inoculum was removed by adsorption with a sterile swab. The mice were examined microscopically for ocular disease using a
scoring system that was described previously (Grau et al., 1989; Brandt et al., 1992). In brief, eyelid inflammation, blepharitis, was scored as follows: (1+) noticeably puffy eyelids, (2+) puffy eyelids with moderate crusting, (3+) eyes wollen, 50% shut with severe crusting, and (4+) eye totally swollen and crusted shut. Vascularization was scored as follows: (1+) 5% of the cornea involved, (2+) <25 to 50% of the cornea involved, and (3+) >50% of the cornea involved. Stromal keratitis was scored as follows: (1+) cloudiness, some iris detail visible, (2+) iris detail obscured, (3+) cornea totally opaque, and (4+) cornea perforated. Ten mice in each group were scored for ocular disease.
Data were reported as the mean disease score for each mouse group on the day of observation, or as the mean peak disease score (MPDS). The MPDS was calculated by averaging the most severe disease score for each animal in a group regardless of the day when the peak score was observed. This model is not reliable for scoring epithelial keratitis because of difficulties in distinguishing virus-induced damage from scarification during infection. Thus, the corneas were not stained with fluorescein, and disease was scored in a masked fashion.
2.4.2. Treatment protocol
A high (GFAHP-HD; 150 μg/ml), medium (GFAHP-MD; 37.5 μg/ml), and low (GFAHP-LD; 3.8 μg/ml) dose of GFAHP, or 1000 μg/ml Acyclovir (ACV, Pusheng BioTech. Co., Wuhan, China) was administered topically in 10 μl saline onto the right cornea. The mice were treated six times per day for the first 3 days, beginning 3–4 h p.i., and four times per day for the next 4 days. Mice receiving six treatments per day were treated at 9:00 a.m., 12:00 p.m., 3:00 p.m., 6:00 p.m., 9:00 p.m., and midnight. Mice receiving four treatments per day were treated at 9:00 a.m., 1:00 p.m., 5:00 p.m., and 9:00 p.m. Ocular disease scores were recorded until day 18 p.i. In control animals, typical dendritic keratitis usually appeared between days 3 and 5 p.i.
2.4.3. Measuring ocular HSV-1 replication
To quantitate virus in the mouse corneas, mice were sacrificed with halothane 24 and 48h p.i. and the right eyeballs were obtained. The eyeballs were homogenized in 4 ml PBS containing 0.2% BSA at 48 ℃ and the homogenate supernatants were prepared as described previously (Grau et al., 1989). The supernatants from two different eyeballs were pooled, and the virus was quantified by plaque assay (Pirano and Brandt, 1999). The results of groups given GFAHP or ACV were compared to those from the untreated groups.
2.5. GFAHP toxicity
2.5.1. Cytotoxicity assay
As a preliminary test of toxicity, replicate wells of uninfected cells were exposed to various concentrations of each substance and incubated for 16–18 h at 37 ℃. Viable cell yield was determined using the MTT reduction assay as previously described (Scubiero et al., 1988). Cytotoxicity was expressed as the 50% cytotoxic concentration (CC50 ) at which a substance inhibited up to 50% of cell growth, as shown by MTT viability 48 h later.
2.5.2. Toxicity in vivo
Uninfected mice were treated with 10 μl 150 μg/ml GFAHP using the protocol described above. Eight hours after the last treatment, the corneas were stained with 1% fluorescein and examined with a cobalt blue light source for evidence of punctate keratopathy, ulceration, or other epithelial defects. A total of 10 mice were examined.
2.6. Stability of GFAHP
Equal volumes of GFAHP in phosphate buffered saline (PBS) were mixed with DMSO ether, or 80% acetone for 15 min and incubated for an additional 15 min at room temperature. For organic solvents, the aqueous layers were removed and the samples were air dried or lyophilized, resuspended at the original concentration (0.1 mg/ml) in DMEM, and tested for antiviral activity. To measure heat stability, GFAHP samples in PBS were boiled for 10 min, cooled to room temperature, and tested for antiviral activity in vitro. To test protease sensitivity, 10 mg/ml GFAHP was incubated with 1.25% trypsinase overnight at 37 ℃. 0.5% Soybean trypsinase inhibitor (STI) was added to inactivate the trypsinase and the samples were tested for antiviral activity in vitro. The percent reduction in antiviral activity was determined by comparing the amount of GFAHP required to reduce viral titers [(untreated/treated)×100]. Control cultures were exposed to a solution containing STI and no GFAHP and STI was found to have no effect (data not shown).
2.7. Statistical analysis
Student’s t-test, analysis of variance (ANOVA), and calculation of correlation coefficients were carried out using SPSS 10. Statistically significant differences were calculated at the 5% significance level using the Fisher’s protected least significant difference (PLSD) test.
3.Results
3.1. Isolation and purification of GFAHP from G. Frondosa
To investigate the antiviral activity of G. frondosa extracts, a protein designated GFAHP was isolated from water-soluble fractions of G. frondosa by ammonium sulfate precipitation and DEAE-cellulose column chromatography. The anti-HSV-1 activity of GFAHP was examined by plaque reduction assay and found to exhibit potent antiviral activity. As shown in Table1, the IC100 for the crude extract against HSV-1 was 512μg/ml. The 40% ammonium sulfate precipitate was found to contain all of the antiviral activity (IC100 , 64μg/ml). The active material was chromatographed on a DEAE-cellulose column and the antiviral activity of each fraction was tested. We got four fractions from the chromatograph, and only the second fraction (total number was fraction 6) had an IC100 of 11μg/ml against HSV-1 KOS (Table 1). Altogether, the protocol resulted in approximately 70× purification with a 0.06% yield. The IC50 value of GFAHP was 4.1μg/ml, and its CC50 value was more than 120μg/ml (Table 2). The therapeutic index (CC50/IC50) of GFAHP was more than 29.3.
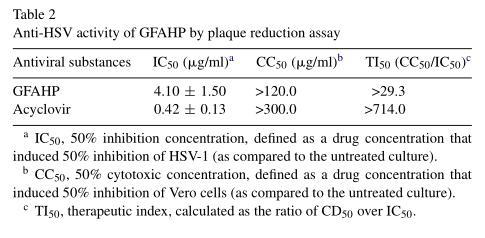
3.2. Possible mechanism of GFAHP antiviral activity in vitro
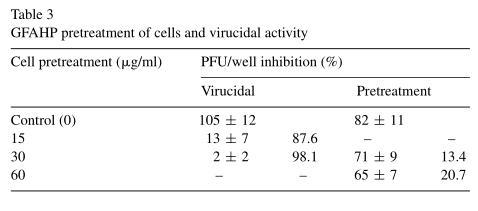
The preventive and virucidal effects of GFAHP, in addition to its effect on HSV-1 attachment and penetration, were determined in order to define its potential mechanism of action. The number of plaques formed by HSV-1in GFAHP pretreated Vero cells was not significantly different from the number formed in untreated Vero cells (Table 3). This suggested that GFAHP may not protect cells from infection. GFAHP was found to significantly decrease viral titer compared with the control group, however, suggesting that it does have virucidal activity. This effect was assumed to be irreversible (Table 3).
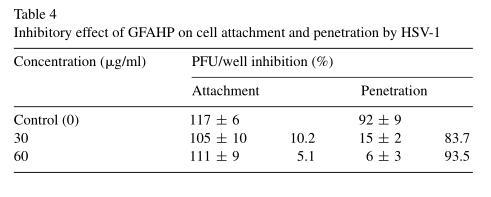
Viruses were allowed to attach to cells in the presence or absence of GFAHP for 3 h. The assay was terminated by removing the virus with PBS (pH 7.4). Treatment with 30 or 60 μg/ml GFAHP was not found to inhibit HSV-1 attachment (Table 4). To examine the effect of GFAHP on penetration, viruses were allowed to attach at 4 ℃ for 2h, a temperature sufficient for attachment but not penetration, after which the temperature was abruptly increased to 37 ℃ to facilitate penetration. 30 μg/ml GFAHP prevented 83.7% HSV-1penetration into cells and 60 μg/ml GFAHP inhibited 93.5% HSV-1 penetration (Table 4). Thus, GFAHP may exert its antiviral activity by preventing HSV penetration into Vero cells.
Based on the above results, GFAHP may have a direct virucidal effect on viral activity in vitro and inhibits viral penetration into cells.
3.3. GFAHP antiviral activity in vivo
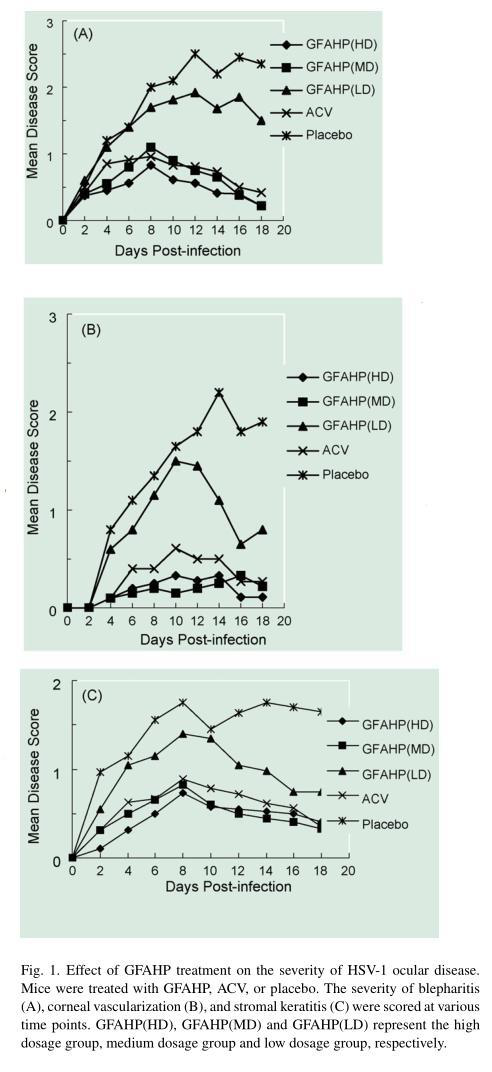
3.3.1. Effect of GFAHP treatment on the development of ocular disease
To determine if GFAHP treatment was effective against ocular disease, mice were infected and treated with ACV, GFAHP, or placebo (Fig. 1A–C). Disease development in the placebo group was similar to that observed in untreated infected mice (data not shown). Blepharitis was evident in all groups by day four, with scores of approximately 1 (Fig. 1A). Blepharitis continued to increase in the placebo and GFAHP-LD treated groups, peaking on day 12 with scores ranging from 1.92 to 2.5. In the GFAHP-HD, MD, and ACV treated groups, blepharitis peaked on day eight with scores of approximately 1, and began to heal soon thereafter. By day 18, when scoring was halted, blepharitis had essentially healed in the GFAHP-HD and MD treatment groups and had nearly healed in the ACV group. In the GFAHP-LD and placebo groups, blepharitis had not healed by this time point.
Corneal neovascularization was evident by day six through eight in all groups (Fig. 1B). In the placebo group, vascularization continued to increase in severity, reaching a score of 2.2 on day 14, when scoring was halted. In the GFAHP-HD, GFAHP-MD, and ACV treatment groups, vascularization also increased, but peaked on day 18 with a score between 0.2 and 0.5, substantially less than that observed in the placebo group. In the group treated with GFAHP-LD, vascularization peaked on day 10 with a score of 1.45 and declined to 0.65 on day 16.
Stromal keratitis was evident by day four and peaked on day eight in all groups (Fig. 1C). In the placebo group, stromal disease increased in severity, reaching a score of 1.75 on day eight, when scoring was halted. In the ACV and GFAHP treated groups, stromal disease was not as severe as in the placebo group. On day 18, the stromal disease scores were 0.4, 0.33, and 0.36 for the GFAHP-HD, GFAHP-MD, and ACV treated groups, respectively, and 1.6 for the placebo group.
3.3.2. Effect of GFAHP on disease incidence
GFAHP treatment had very little effect on the incidence of blepharitis. In the placebo, ACV, GFAHP-LD, and GFAHP-MD treated groups, 100% of the mice developed disease. In the GFAHP-HD treated groups, all but two of the mice developed blepharitis (77.8%). Peak incidence was reached between days 4 and 8 by all groups. The incidence declined after day 8 in the GFAHP-HD treated group, and reached 22.2% by day 18. GFAHP also reduced the incidence of vascularization from 100% in the placebo group to between 22% and 56% in the GFAHP-HD and GFAHP-MD treated groups. Treatment further reduced the incidence of stromal disease from 100% in the placebo group to approximately 70% in the GFAHP-HD and GFAHP-MD treated groups. In the GFAHP-HD treated groups, only two mice (22%) had permanent corneal clouding on day 16 (data not shown).
3.3.3. MPDSs
Mean peak disease score (MPDS) is used to compare the disease scores of multiple groups (Grau et al., 1989.). Statistical analysis of the MPDSs revealed that GFAHP-LD did not significantly reduce blepharitis, vascularization, or stromal disease, with p values of 0.08, 0.12, and 0.09, respectively. Treatment with GFAHP-HD, GFAHP-MD, and ACV had a significant effect on blepharitis, however (p<0.005). GFAHP-HD and GFAHP-MD also significantly reduced the MPDSs for vascularization and stromal disease (p<0.01) (data not shown).
3.3.4. Antiviral effect of GFAHP-I on HSV-1 replication in the mouse cornea
We quantitated the amount of HSV-1 recovered from the eyeballs of mice treated with GFAHP-HD or ACV after virus inoculation. The mean virus titer in GFAHP-HD treated mice was 12.7% of that observed in the control group (p<0.01) 24 h p.i. After 48 h, the mean virus yield in the GFAHP-HD group was 23.5% of that observed in the control group (p<0.01). The virus yield after GFAHP-HD treatment was significantly lower than after ACV treatment (p<0.05). ACV treatment also suppressed viral replication in the cornea, reaching 40.38% and 53.5% of the control groups at 12 and 24 h, respectively (p<0.05 versus control group) (Fig. 2).
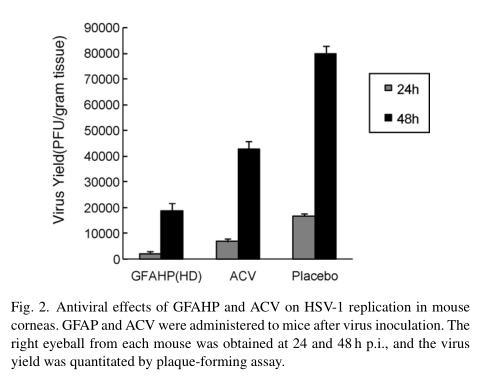
3.3.5. Histopathology of infected corneas
Corneal sections from day 18 infected mice were substantially thicker and edematous after placebo than GFAHP-HD treatment. The epithelium showed more evidence of necrosis and polymorphonuclear leukocyte infiltration after the placebo, and intranuclear inclusion bodies usually associated with HSV-1 infection were present. There was also a stromal infiltrate consisting of polymorphonuclear leukocytes and epitheloid cells. The corneal stroma was vascularized, and there were typical signs of both acute and chronic interstitial keratitis typical of HSV-1 corneal infection. The corneas from infected mice in the GFAHP-HD treatment group were almost normal, showing no evidence of inflammation, necrosis, or cells containing inclusion bodies (data not shown).
3.4. GFAHP toxicity in vivo
To determine if GFAHP was toxic in vivo, uninfected mice were treated with either placebo or GFAHP and at the end of the treatment period the corneas were stained with fluorescein and examined for epithelial defects. No evidence of corneal damage such as punctate keratopathy was observed in either placebo or GFAHP treated mice (data not shown). Importantly, injured corneas of mice treated with and without GFAHP healed in a similar period of time, and corneal ulceration produced by needle scratching healed within 2 days regardless of GFAHP treatment. Histopathological analysis of the corneas revealed no discernible differences between GFAHP treated and untreated corneas (data not shown).
3.5. Identification of GFAHP
GFAHP was resuspended at 0.1mg/ml in distilled water and treated with lipid solvents, ether, DMSO, acetone, ortrypsinase, to determine the nature of its antiviral activity. Ether, acetone and DMSO had no effect on the antiviral activity of GFAHP. Treatment with trypsinase digestion reduced GFAHP activity to 27% after overnight incubation. These results illustrate that GFAHP is likely to be a protein or to have a proteinaceous component. Boiling GFAHP for 10min did not reduce its antiviral activity.
GFAHP was isolated from the 40% ammonium sulfate precipitate of G.frondosa and its structure was identified using both physical and chemical analyses. Analysis of the second peak (fraction 6-GFAHP) with SDS-PAGE indicated that GFAHP contained one band with a molecular weight of approximately 29.5kDa (Fig. 3). A piece of the filter containing the 29.5kDa protein was then cut from the SDS-PAGE gel, and sequenced by MALDI-TOF-MS and nano ESI-MS/MS. The N-terminal sequence of GFAHP consisted of an 11 amino acid peptide, NH2-REQDNAPCGLN-COOH. The data was processed using Mascot software and a database search was performed with the MS/MS ion on the Mascot engine. The sequences were not found to match any known amino acid sequences. The molecular weight of this protein from the MALDI-TOF-MS was 29.529kDa.
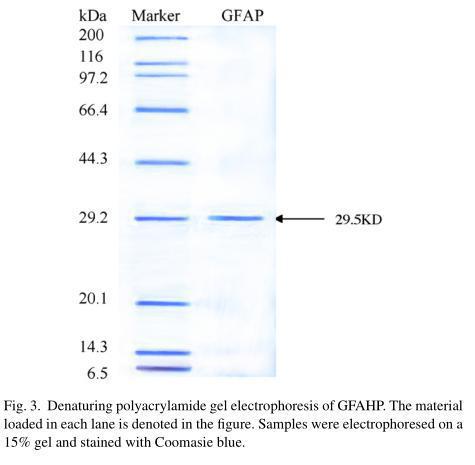
4.Discussion
The present study described the isolation, partial structural characterization, and in vitro and in vivo activity of an anti-HSV-1 substance isolated from the edible mushroom, G. Frondosa. GFAHP was isolated from water-soluble fractions of G. frondosa by ammonium sulfate precipitation and DEAE-cellulose column chromatography. Based on the anti-HSV-1 activity, the 40% ammonium sulfate precipitate had kept all of the antiviral activity. The active material was isolated and purified through DEAE-cellulose chromatographic column, and finally we got the antivirus activation fraction.
The IC50 value of GFAHP was 4.1 and 0.42μg/ml was for ACV. We investigated the mechanism of GFAHP antiviral activity in vitro by evaluating its ability to prevent infection and directly inhibit HSV-1 by preventing replication and cell adsorption and penetration. We found that GFAHP had direct virucidal effect on HSV, and inhibited viral entry, but could not prevent virus infection, or inhibit adsorption and replication. In a previous study, we reported that the D-Fraction (GF-D) extracted from G. frondosa had a significant inhibitory effect on hepatitis B virus (HBV) in HepG22.2.15 cells, and may also inhibit HBV replication (Gu et al., 2006).
When mice were infected with HSV-1 KOS and treated with various concentrations of GFAHP, the severity of ocular disease, as measured by scoring blepharitis, corneal vascularization, and stromal keratitis, was reduced in a dose-dependent manner. With the increase on GFAHP concentration, these verity of blepharitis, vascularization and stromal disease was reduced. The medium and high concentrations of GFAHP significantly reduced the severity of these diseases compared with the control and low GFAHP groups.
With the exception of ACV, many currently available nucleoside analog antiviral agents exhibit toxicity even when used topically (Squires, 2001). In the present study we showed that application of the high dose of GFAHP (150 μg/ml) for seven consecutive days (34 treatments) did not cause conjunctivitis or blepharitis, indicating that this protein has little if any toxicity, at least when applied topically. Histopathological examination of treated corneas also showed no evidence that the high dose of GFAHP is toxic to cells.
Active GFAHP was heat stable and partially sensitive to trypsinase digestion. Trypsinase was able to reduce GFAHP activity by 73% but unable to completely eliminate it. These findings illustrated that GFAHP is likely to be a protein or to have a proteinaceous component. Analysis using gel electrophoresis, MALDI-TOF-MS, and nano ESI-MS/MS suggested that it had a molecular weight of 29.5 kDa, and an N-terminal sequence with 11 aminoacidpeptides, NH2 -REQDNAPCGLN-COOH. The GFAHP N-terminus was used to search the SWISS-PORT and NCBIInr databases using the Mascot search engine (http://www.matrixscience.co.uk) with a tolerance of ±0.1Da and one missed cleavage site. Three proteins with high similarly were chosen to analyze. The hypothetical protein, TTHERM 00324530, shared 87% similarity with GFAHP (7/8), and the 632nd amino acid in its N-terminus matched the first amino acid of GFAHP, however the function of this protein is unknown. One serine proteinase shared 82% similarity with the GFAHP N-terminus (9/11), and the 92nd amino acid in its N-terminus matched the first amino acid of GFAHP. Serine proteinases are involved in many processes of critical medical significance and are often involved in severe pathophysiological diseases (Czapinska and Otlewski, 1999). However, there is no report about that this protease has antiviral activity. Hypothetical protein, TTHERM 00637630 had 87% (7/8) similarity with GFAHP and its 206th amino acid matched with the 4th amino acid of GFAHP, however the function of this protein is also unknown. Thus, it is likely that GFAHP is a novel protein with antiviral activity.
A number of antiviral proteins have been described. Three different proteins with similar antiviral activity have been isolated from pokeweed, Phytalacca Americana (Hartley et al., 1991; Irvin and Uckun, 1992; Wang et al., 2006). Trichosanthes kirilowii contains a 29-kDa material that inhibits HIV (Lee-Huang et al., 1991). A protein of 10,425Da purified from the edible mushroom, Rozites caperata, was shown to inhibit HSV-1 and -2 replication (Pirano and Brandt, 1999). A proteinaceous material with broad spectrum antiviral activity was isolated from the cactus, Opuntia streptocantha (Ahmad et al., 1996), and lectins are known to have activity against many different viruses (Ooi et al., 2004; Charan et al., 2000; Ye et al., 2001). A protein from the edible mushroom, R. caperata, has antiviral activity against HSV, varicella zoster, IVA, and respiratory syncytial virus (Pirano and Brandt, 1999). Velutin, a ribosome inactivating protein from Flammulina velutipes, is shown to inhibit HIV-1 reverse transcriptase (Wang and Ng, 2001), and a protein from G. frondosa inhibits tobacco mosaic virus infection (Chen et al., 2004). Recently, Pirano (2006) observed on the characterization and antiviral properties of a novel antiviral protein, RC 28, isolated from the mushroom Rozites caperatus. It is unlikely that GFAHP is related to a previously reported antiviral protein since its amino acid sequence did not match proteins with known antiviral activity. Thus, it is probable that GFAHP is a novel structure.
In summary, we have isolated and partially characterized a novel protein from the edible mushroom, G. frondosa, with potent antiviral activity. The antiviral substance, GFAHP which has a favorable IC50 value of 4.1 μg/ml, is active against HSV-1 both in vitro and in vivo. Confirming the structure and determining the mechanism of GFAHP antiviral function should aid development of a new antiviral and help to identify novel features of the viral life cycle.
References
Adachi, Y., Okazaki, M., Ohno, N., Yadomae, T., 1994. Enhancement of cytokine production by
macrophages stimulated with (1→3)-β-glucan, grifolan, isolated from Grifola frondosa. Biol.
Pharm. Bull. 17, 1554–1560.
Ahmad, A., Davies, J., Randall, S., Skinner, G.R.B., 1996. Antiviral properties of extract of
Opuntia streptacantha. Antivir. Res. 30, 75–85.
Akihisa, T., Franzblau, S.G., Tokuda, H., Tagata, M., Ukiya, M., Matsuzawa, T., Metori, K.,
Kimura, Y., Suzuki, T., Yasukawa, K., 2005. Antitubercular activity and inhibitory effect
Epstein-Barr virus activation of sterols and polyisoprenepolyols from an edible mushroom,
Hypsizigus marmoreus. Biol. Pharm. Bull. 28, 1117–1119.
Aoki, M., Tan, M., Fukushima, A., Hieda, T., Mikaml, Y., 1993. Antiviral substances with septimic
effects produced by basidiomycetes such as Fomes fomentarius. Biosci. Biotechol. Biochem.
57, 178–182.
Borchers, A.T., Stern, J.S., Hackman, R.M., 1999. Mushrooms, tumors and immunity. Proc. Soc.
Exp. Biol. Med. 221, 281–293.
Brandt, C.R., Coakley, L.M., Grau, D.R., 1992. A murine model of herpes simplex virus-induced
ocular disease for antiviral drug testing. J. Virol. Methods 36, 209–222.
Charan, R.D., Munro, M., O’Keefe, B.R., Sowder, R., McKee, T., Currens, M., Pannell, L.K.,
Boyd, M.R., 2000. Isolation and characterization of Myrianthus holstii lectin, a potent HIV-1
inhibitory protein from the plant Myrianthus holstii(1). J. Nat. Prod. 63, 1170–1174.
Chen, N., Wu, Z.J., Lin, Q.Y., Ie, L.H., 2004. Purification and partial characterization of a protein
inhibitor of tobacco mosaic virus infection from Grifola frondosa. Prog. Biochem. Grifola
Biophys. 31, 283–286.
Chiang, L.C., Chiang, W., Chang, M.Y., Ng, L.T., Lin, C.C., 2002. Antiviral activity of Plantago
major extracts and related compounds in vitro. Antivir. Res. 55, 53–62.
Collins, R.A., Ng,T.B., 1997. Polysaccharopeptide from coriolus versicolor has potential for use
against human immunodeficiency virus type 1 infection. Life Sci. 60, PL383–PL387.
Cui, F.J., Tao, W.Y., Xu, Z.H., Guo, W.J., Xu, H.Y., Ao, Z.H., Jin, J., Wei, Y.Q., 2007. Structural
analysis of anti-tumor heteropolysaccharide GFPS1b from the cultured mycelia of Grifola
frondosa GF9801. Bioresource Technol. 98, 395–401.
Czapinska, H., Otlewski, J., 1999. Structural and energetic determinants of the S1-site specificity
in serine proteases. Eur. J. Biochem. 260, 571–595.
De Logu, A., Loy, G., Pellerano, L.M., Bonsignore, L., Schivo, M.L., 2000. Inactivation of HSV-1
and HSV-2 and prevention of cell-to-cell virus spread by Santolina insularis essential oil.
Antivir. Res. 48, 177–185.
Grau, D.R., Visalli, R.J., Brandt, C.R., 1989. Herpes simplex virus stromal keratitis is not
titer-dependent and does not correlate with neurovirulence. Invest Ophthalmol. Vis. Sci. 30,
2474–2480.
Gu, C.Q., Li, J.W., Chao, F.H., 2006. Inhibition of hepatitis B virus by D-Fraction from Grifola
frondosa: synergistic effect of combination with interferon-α in HepG2 2.2.15. Antivir. Res.
72, 162–165.
Hartley, M.R., Leguame, G., Osborn, R., Chen, Z., Lord, J.M., 1991. Single chain ribosome
inactivating proteins from plants depurinate Escherichia coli 23S ribosomal RNA. FEBS
Lett. 290, 65–68.
Hsieh, C., Liu, C.J., Tseng, M.H., Lo, C.T., Yang, Y.C., 2006. Effect of olive oil on the production
of mycelial biomass and polysaccharides of Grifola frondosa under high oxygen
concentration aeration. Enzyme Microb. Tech. 39, 434–439.
Inoue, A., Kodama, N., Nanba, H., 2002. Effect of maitake (Grifola frondosa) D-Fraction on the
control of the T lymph node Th-1/Th-2 proportion. Biol. Pharm. Bull. 25, 536–540.
Irvin, J.D., Uckun, F.M., 1992. Pokeweed antiviral protein ribosome inactivation and therapeutic
applications. Pharmacol. Ther. 55, 279–302.
Kodama, N., Komuta, K., Sakai, N., Nanba, H., 2002. Effects of D-Fraction, a polysaccharide
from Grifola frondosa on tumor growth involve activation of NK cells. Biol. Pharm. Bull. 25,
1647–1650.
Kodama, N., Asakawa, A., Inui, A., Masuda, Y., Nanba, H., 2005. Enhancement of cytotoxicity of
NK cells by D-Fraction, a polysaccharide from Grifola frondosa. Oncol. Rep. 13, 497–502.
Lee, B.C., Baea, J.T., Pyoa, H.B., Choe, T.B., Kimc, S.W., Hwang, H.J., Yun, J.W., 2003.
Biological activities of the polysaccharides produced from submerged culture of the edible
basidiomycete Grifola frondosa. Enzyme Microb. Tech. 32, 574–581.
Lee-Huang,S.,Huang,P.L.,Jung,H.F.,Li,B.-Q.,Huang,P.L.,Huang,P.,Huan, H.I., Chen, H.-C., 1991.
TAP29: an antihuman immunodeficiency virus protein from Trichosanthus kirilowii is
non-toxic to intact cells. Proc. Natl. Acad. Sci. U.S.A. 88, 6570–6574.
Lee, B.C., Baea, J.T., Pyoa, H.B., Choe, T.B., Kimc, S.W., Hwang, H.J., Yun, J.W., 2004.
Submerged culture conditions for the production of mycelial biomass and exopolysaccharides
by the edible basidiomycete Grifola frondosa. Enzyme Microb. Tech. 35, 369–376.
Mayell, M., 2001. Maitake extracts and their therapeutic potential. Altern. Med. Rev. 6, 48–60.
Mentel, R., Meinsen, D., Pilgrim, H., Herrmann, B., Lindequist, U., 1994. In vitro antiviral effect
of extracts of Kuehneromyces mutabilis on influenza virus. Pharmazie 49, 859–860.
Mothana, R., Awadh, N., Jansen, R., Wegner, U., Mentel, R., Lindequist, U., 2003. Antiviral
lanostanoid triterpenes from the fungus Ganoderma pfeifferi BRES. Fitoterapia 74, 177–180.
Nanba, H., Kodama, N., Schar, D., Turner, D., 2000. Effects of maitake (Grifola frondosa) glucan
in HIV-infected patients. Mycoscience 41, 293–295.
Nie, X., Shi, B., Ding, Y., Tao, W., 2006. Preparation of a chemically sulfated polysaccharide
derived from Grifola frondosa and its potential biological activities. Int. J. Biol.
Macromolecules 39, 228–233.
Ooi, L.S., Sun, S.S., Ooi, V.E., 2004. Purification and characterization of a new antiviral protein
from the leaves of Pandanus amaryllifolius (Pandanaceae). Int. J. Biochem. Cell B 36,
1440–1446.
Pirano, F., Brandt, C.R., 1999. Isolation and partial characterization of an antiviral,RC-183, from
the edible mushroom Rozites caperata. Antivir. Res. 43, 67–78.
Pirano, F., 2006. Emerging antiviral drugs from medicinal mushrooms. Int. J. Med. Mushrooms. 8,
101–114.
Rosenthal, K.S., Peres, R., 1985. Inhibition of herpes simplex virus type 2 penetration by
cytochalasins D and B. J. Gen. Virol. 66, 1601–1605.
Sarkar, S., Koga, J., Whitley, R.J., Chatterjee, S., 1993. Antiviral effect of the extract of culture
medium of Lentinas edodes mycelia on the replication of herpes simplex virus type 1. V
Antivir. Res. 20, 293–303.
Scubiero, D.A., Shoemaker, R.H., Paull, K.D., Monks, A., Tierney, S., Nofziger, T.H., Currens,
M.J., Seniff, D., Boyd, M.R., 1988. Evaluation of a soluble tetrazolium:formazan assay for
cell growth and drug sensitivity in culture using human and other tumor cell lines. Cancer
Res. 48, 4827–4833.
Shi,B.J.,Nie,X.H.,Chen,L.Z.,Liu,Y.L.,Tao,W.Y.,2006.Anticanceractivities of a chemically sulfated
polysaccharide obtained from Grifola frondosa and its combination with 5-Fluorouracil
against human gastric carcinoma cells. Carbohydr. Polym. 68, 687–692.
Sorimachi, K., Akir, N., Yamazaki, S., Toda, S., Yasumura, Y., 1990. Anti-viral activity of
water-solubilized lignin derivatives in vitro. Agric. Biol. Chem. 54, 1337–1339.
Squires, K.E., 2001. An introduction to nucleoside and nucleotide analogues. Antivir Ther. 6
(Suppl. 3), 1–14.
Visalli, R.J., Brandt, C.R., 1993. The HSV-1 UL45 18-kDa gene product is a true late protein and
a component of the virion. Virus Res. 29, 167–178.
Wang, H., Xia, Q., Li, P., He, J., Wang, J., Zhang, X., 2006. N-terminal acetylation
characterization of tumor protein D52 by nano electrospray ionization tandem mass
spectrometry. Chin. J. Anal. Chem. 34, 321–324.
Wang, H.X., Ng, T.B., 2001. Isolation and characterization of velutin, a novel
low-molecular-weight ribosome-inactivating protein from winter mushroom (Flammulina
velutipes) fruiting bodies. Life Sci. 68, 2151–2158.
Ye, X. Y., Ng, T.B., Tsang, P. W., Wang, J., 2001. Isolation of a homodimeric lectin with antifungal
and antiviral activities from red kidney bean (Phaseolus vulgaris) seeds. J. Protein Chem. 20,
367–375.
本文部分内容来源网络,如有侵权请联系删除,谢谢。
编辑:中润世源
