口服舞茸子实体提取物表现出的抗肿瘤活性研究
日期:2019-07-31 / 人气:
声明:本文摘自Chem. Pharm. Bull.,全称Chemical and Pharmaceutical Bulletin ,中文名《化学与药学通报》,由日本药学院(PSJ)出版,1953年创刊,全年12期,发表生物分析化学、生物化学、药理学、毒理学和生物药学方面的研究论文及报告,用英文出版。为方便广大读者阅读,对部分内容进行了中文翻译,译文仅供参考,如有不足之处,敬请指正。
【参考译文】
Antitumor Activity Exhibited by Orally Administered Extract from Fruit Body of Grifola frondosa (Maitake)
口服舞茸子实体提取物表现出的抗肿瘤活性研究
从舞茸子实体提取的酸不溶性、碱溶性、热水溶性高分子聚合物(一种约含30%蛋白的多糖;舞茸蛋白多糖)在小鼠口服试验中表现出抗同种肿瘤和同系肿瘤活性。Winn的实验结果显示出了舞茸蛋白多糖给药对肿瘤生长的抑制作用。这一事实表明舞茸蛋白多糖可激活荷瘤状态的免疫系统。因此,证明了舞茸蛋白多糖的抗肿瘤细胞活性与免疫反应是相互关联的。巨噬细胞或T细胞的细胞溶解活性和白细胞介素的生产能力所展现出的抗原特异性细胞毒性有所提高。此外,舞茸蛋白多糖被发现可以增强与抑制肿瘤生长相关的迟发型超敏反应。
关键词:舞茸;抗肿瘤活性;β-葡聚糖(舞茸蛋白多糖);迟发型超敏反应;口服给药
【论文原文】
Chem. Pharm. Bull.
36(5), 1988, 1819-1827
Antitumor Activity Exhibited by Orally Administered Extract from Fruit
Body of Grifola frondosa (Maitake)
IKUKO HISHIDA,HIROAKI NANBA* and HISATORA KURODA
Laboratory of Microbiology, Kobe Women's College of Pharmacy,
Motoyama, Higashinada, Kobe 658, Japan
(Received September 14, 1987)
The acid-insoluble, alkali-soluble,hot-water-extractable polymer(a polysaccharide containing approximately 30% of protein; D-fraction) obtained from the fruit bodies of Grifola frondosa
(maitake) exhibited antitumor activities against allogenic and syngeneic tumors on oral administration to mice. Winn assay conducted to examine the tumor growth-suppressing effect revealed a complete inhibition of the tumor by the oral administration of D-fraction. This fact indicates that stimulation of the immune response system triggered by the tumor-bearing state is activated by D-fraction. Consequently, the activity of D-fraction on cells associated with immune response was examined. The cytolytic activity and interleukin-1 productivity pf macrophages or T cells which exhibit antigen-specific cytotoxicity were enhanced. D-fraction was found to potentiate the delayed-type hypersensitivity response which is associated with tumor growth suppression.
Keywords——Grifola frondosa ; antitumor activity; β-glucan(D-fraction); delayed-type hypersensitivity (DTH); oral administration
Many antitumor-active polysaccharides derived from basidiomycetes have been reported, including lentitan, shizophyllan, PSK and others, most of which principally consist of similar β-glucans.Most of these polysaccharides, excluding PSK, exhibit antitumor activity via the activation of host immune systems when administered intravenously or intraperitoneally. However, they have been reported to be ineffective when given orally. The authors have isolated from the fruit body of Grifola frondosa(maitake), a β-glucan(MT-2) which carries many 1,3-linked oligosaccharied side chains branching from the 1,6-linked main chain. The authors reported previously1,2) that MT-2 exhibited antitumor activity by stimulating the immune system of mice, when administered intraperitoneally. Miyazaki3) and Mizuno et al.4) obtained a lentinan-like polysaccharide from this fruit bodies and reported that was inactive when administered orally , whereas an antitumor effect was exhibited when it was administered intraperitoneally or intravenously. In contrast, the author5,6) have observed that the immune systems in mice were stimulated, leading to the regression of tumors, when powdered fruit body of Lentinus edodes(shiitake) was orally administered to mice. Moreover, Ohkuma et al.,7) reported that stimulation of the immune system occured when a protein-poly-saccharide complex extracted with hot water from Flammulina velutipes was administered orally to tumor-bearing mice. In this context, we have examined the antitumor activity of extracts from the bodies of maitake.
Maitake and Methods
(1) Preparation of Extractable Maitake——The process used for extracting materials from the powdered fruit body of Grifola frondosa ,supplied by the Mushroom Research Institute of Japan(Kiryu Gumma), was based on the described by Chihara er al. for the extraction of lentinan from shiitake.8) As shown in Fig. 1, the powder (600 g) was mixed with 3000ml of deionized water, and heated at 100 ℃ for 10 h. After centrifugation at 6000 rpm for 10 min, the supernatant was used as pre-A fraction. An equal amount of 99% EtOH was added to this pre-A fraction and left at dried fruit bodies of G. Frondosa(200g)
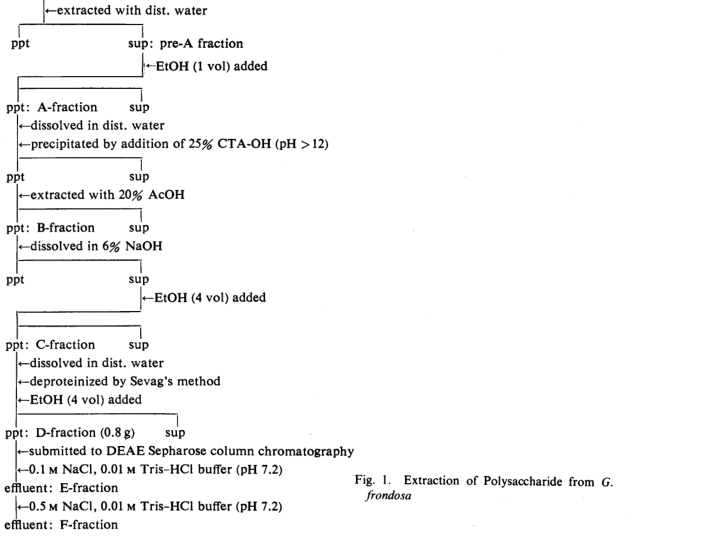
4℃ for 24 h. The precipitate A-fraction was separated and dissolved in deionized water. Then, 25%(CTA-OH) (cetyltrimethylammonium hydroxide) was added dropwise with stirring until the pH exceeded 12 and the mixture was allowed to stand at 4℃ for 12h to give a precipitated. This pellet was treated with 20-50 % acetic acid to remove the acid-soluble fraction, and the residual fraction was obtained as B-fraction. This fraction was treated with 6% NaOH to separate the alkali-soluble fraction. This solution was combined with 4 volumes of EtOH to obtain the precipitate polymer. This pellet(C-fraction) was dissolved in an equal amount of distilled water and deproteinized with CHCl3-MtOH(9:1). The solution was again treated with 4 volumes of EtOH. The obtained precipitate pellet (D-fraction) was a light brown and amorphous power, which was positive in the anthrone test, and contained approximately 30% protein as determine by Lowry's method. The D-fraction was charged on a diethylaminoethyl(DEAE)-Sepharose column (5×40cm), and eluted with 0.01 M Tris-HCl buffer containing 0.1 or 0.5 M NaCl (pH 7.2) to give E-fraction and F-fraction successively.
(2) Mice and Tumors-Sarcoma-180 was transplanted into ICR mice, MM-46 carcinoma and MH-134 hepatoma into C3H/HeN (C3H) mice, I MC carcinoma into CDF1 mice and B-16 ,melanoma into C57BL/6 mice.
(3) Tumor Growth-Inhibitory Effect-Tumor cells (1×106) were implanted subcutaneously in the right axillary region of mice, and from 24h after administration, 0.5 ml of each fraction, adjusted to a sugar concentration of 3.0mg/ml, was orally administered on alternate days. After the completion of the administration, the solid tumors were extirpated and weighed to obtain the tumor growth inhibition rate (TIR), which was calculated according to the previous pater.1)
(4) Winn Assay9)-MM-46 tumor cells (1×106) were inoculated in the axillary region, and from 24h after inoculation, 0.5 ml of D-fraction, adjusted to a sugar concentration of 1.5mg/ml, was orally administered 10 times on alternate days. After the completion of the administration, the spleen was extirpated from these D-fraction-administered mice (hereafter termed simply D-mice). The obtained 5×106 spleen cells were mixed with 1×106 MM-46 Tumor cells. The mixed cell suspension was implanted into the right axillary region of normal C3H mice (recipients), and after 14 d, solid tumors were extirpated and weighed. On the other hand, two experimental systems were employed to compare tumor growth inhibition in D-mice. In the first, the tumor was extirpated from C3H mice inoculated with a mixture of MM-46 tumor cells and the spleen cells of MM-46 tumor-bearing C3H mice which had been given saline orally. In the second, the grpwing tumor was isolated from C3H mice implanted with only MM-46 tumor cells as control mice.
(5) Preparation of Macrophages(MΦ)-Five-week-old C3H (or CDF1) male mice were killed by dislocating cervical vertebrae and peritoneal cells were obtained by washing with Hanks' solution. After centrifugation at 1200rpm for 5 min, the precipitated cells were collected and adjusted with RPMI-1640 medium to 1×105 cells/ml. These cells were incubated for 45 min at 37℃ in an atmosphere containing 5% CO2 gas, and non-adherent cells were eliminated by washing with Hanks' solution. MΦ which were adsorbed selectively on the plate wall were collected.
(6) Preparation of Whole Spleen Cells-The extirpated spleen from C3H male mice (5 weeks old) was washed and disintegrated sufficiently in RPMI 1640 medium. Then, the cells were passed through a 40-mesh stainless steel screen. The cells were collected by centrifugation at 1200 rpm for 10 min and 0.2 ml of a 10-fold dilution of Eagle's MEM was added to lyse contaminating erythrocytes hypotonically for 10 s, then 2 ml of Eagle's MEM (2-fold concentration) was added promptly and mixed. The whole was centrifuged at 1200 rpm foe 10 min, and the obtained cells were adjusted to 1×107 cells/ml and used as the whole spleen cell suspension.
(7) Preparation of T Cells - Whole spleen cell suspension was charged on a nylon wool column (1×15 cm) moistened with RPMI-1640 medium. After incubation for 60 min at 37℃ in a humidified atmosphere of 5% CO2 in air, the T cells were collected by centrifugation at 1200 rpm for 5 min.
(8) Preparation of Lyt-1+ T Cells (10)-Anti-Lyt-2.1 monoclonal antibody (50 μl) was mixed with 1×106 T cell/ml. After incubation at 37℃ for 60 min in an atmosphere of 5% CO2, in air, the suspension was washed with RPMI-1640 medium by centrifugation. The cells obtained were incubated with 50 μl of Low·Tox-M rabbit complement for 60 min at 37 ℃. Then cells obtained by subsequent centrifugation were stained with trypan blue,and a preparation containing 1×106 viable cells was made.
(9) Labeling of the Target Cells- As described in the previous paper,2) 2×106 tumor cells were mixed 50 μl of 40 μCi/ml 3H-uridine and after incubation for 60 min at 37 ℃ in an atmosphere containing 5% CO2 gas, free 3H-uridine was sufficiently rinsed out with RPMI-1640 medium. The same medium was further added to prepare a suspension containing 1×104 cells/ml.
(10) Cytotoxicity Test -The effector cells obtained from D-mice were suspended in RPMI-1640 medium(1×105 cells/ml), to which were added labeled tumor cells to make the E/T ratio=10:1 or 50:1. The cell suspension was cultured at 37 ℃ for a predetermined period of time and after centrifugation, 1ml of the supernatant was mixed with 10 ml of Univagel II reagent (Nakarai Chemicals Co.). The radioactivity of the obtained suspension was measured with an Aloka LSC-700 scintillation counter.
(11) Preparation of Solution Containing Interleukin-I(IL-1)11)-MΦ obtained from D-mice were adjusted to 1×105/ml with RPMI-1640 medium which did not contain serum, and 2 ml of this suspension was placed on a 12-well flat plate. After incubation for 72 h at 37 ℃ in an atmosphere of 5% CO2 in air, the supernatant was filtered through a 0.45 nm Millipore filter for sterilization and stored at -80 ℃. This was thawed as required.
(12) Measurement of Lymphocyte Growth Reaction11) -The thymus was extirpated from normal male C3H mice which had been killed by bleeding from the femoral vein and carotid artery. Cells were teased out in RPMI-1640 medium and after being passed through an 80 mesh sieve, were used as the cell suspension. The suspension was adjusted to 1×107 /ml with RPMI-1640 medium containing 0.7 mg/ml peanut lectin coagulant. After incubation for 45 min, 5 ml of 20% fetal bovine serum (FBS)-added RPMI-1640 medium was overlaid, and after further incubation for 90 min, PNA-noncoagulated cells were collected. These cells were washed with RPMI-1640 medium not containing serum, and cells were resuspended in 2 ml of galactose solution (2.7g of D-galactose, 2 ml of FBS, 15 ml of MEM medium, 3ml of 20mM HEPES and 75 ml of distilled H20). After 10 min, sufficient rinsing was conducted with RPMI-1640 medium, and the suspension were adjusted to 5×106 cells/ml with RPMI-1640 medium and divided into 0.1ml aliquots on a 96-well flat plate (Corning Glass Works). IL-1 solution (0.05ml) obtained as described (11) was added at two dilutions with 0.05ml of RPMI-1640 medium or 0.05 ml of PHA-P at 10μg/ml. This mixture was incubated for 73h. At 8h before the end of incubation, 0.1μCi of 3H-thymidine was added and mixed by shaking the plate. Then 100μl of this reaction fluid was filtered through a Millipore filter, which was dried after being washed with 1.5ml of 5% trichloroacetic acid (TCA) solution. The radioactivity was measured(see preceding section(10)).
Results
As a preliminary test, after implantation of an allogeneic Sarcoma-180 tumor into ICR mice, each extracted fraction was administered, in an amount of 1.5mg sugar content, orally 10 times on alternate days. On the 12th day after the completion of the administration, solid tumors were extirpated and weighed to obtain the TIR. When D-fraction was administered 58% TIR was obtained but practically no antitumor effect was observed with E- or F-fraction (obtained by the passage of D-fraction through an ion-exchange column). Therefore, the authors deal exclusively with D-fraction in this paper. The result is shown in Table I. The D-fraction at doses ranging from 0.2 to 1.5 mg of the sugar was administered to determine the optimal dose and the result is given in Table II. While each of these doses was significantly effective at a probability level of 0.01, no significant difference was observed between these doses. However, intragroup variance was the least at the 0.75 mg dose(25 μg/g of mouse body weight). Therefore this dose of D-fraction was used in this experiment.
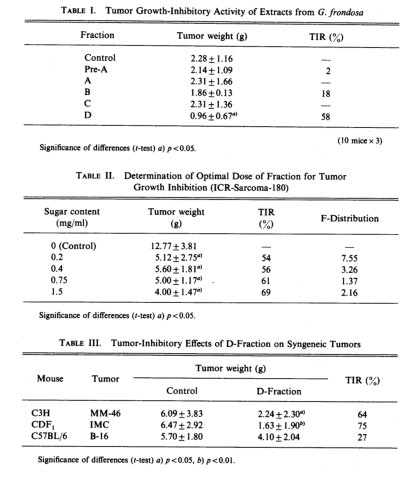
Table III indicates the growth-inhibitory effect of the D-fraction against some syngeneic tumors. MM-46 carcinoma in C3H mouse and IMC carcinoma in CDF1 regressed after oral administration of D-fraction under these conditions (TIR,64% and 75%), while the tumor growth inhibition was as low as 27% in the case of B-16 melanoma in C57BL/6 mouse. Winn assay was conducted to examine whether immune systems were associated with the manifestation of these antitumor effects. MM-46 carcinoma tumor cells were subcutaneously implanted in C3H mice, and D-fraction was orally administered according to the previous procedures. Thus mice in which the tumor growth was suppressed 60 to 70% were termed D-fraction. A spleen cell suspension obtained from these. D-mice or MM-46 tumor-bearing mice was mixed with MM-46 carcinoma tumor cells and the cell suspension was inoculated into normal C3H mice. After breeding the mice for 14d, the extirpated tumors were weighed. Table IV shows the results. While significant regression was observed in tumors obtained from the mice implanted with mixture of tumor cells and spleen cells obtained frpm tumor-bearing mice, complete tumor destruction was observed in mice inoculated with mixture of tumor cells and spleen cells obtained from D-mice. These results suggest that the activity of the immune response system which had been induced by the implantation of the tumor was further stimulated by the administration of D-fraction.
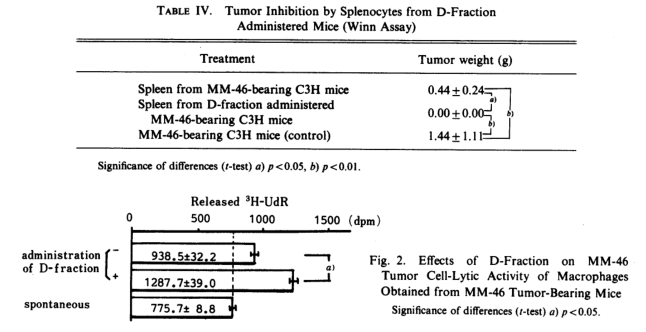
Therefore, we examined how the cells of the immune system were activated by D-fraction in an in vitro system. First, intraperitoneal MΦ obtained from D-mice or from tumor-bearing mice not given D-fraction were mixed with 3H-uridine-labeled tumor cells(at an E/T ratio=1/50) and after incubation for 48h, the released radioactivity in the medium was measured. As shown in Fig. 2, MΦ obtained from normal mice did not exhibit any cytotoxicity to tumor cells, but MΦ obtained from MM-46 tumor-bearing mice did show cytotoxicity . Administration of D-fraction further potentiated this activity.
Next, the action of D-fraction against cytotoxic T cells (Tc cells), which show antigen-specific, was examined. The T cell suspension which was obtained from spleen cells of D-mice or control mice by passage a nylon-wool colum was mixed with 3H-uridine-labeled MM-46 or MH 134 tumor cells as syngeneic timor cells and IMC tumor cells as allogeneic tumor cells, (E/T=1/10), After incubation for 4h, the radioactivity released into the suspernatant was measured; the results are summarized in Fig. 3. The activity of Tc cells obtained from D-mice, the MM-46 tumor of which had regressed, exhibited approximately 3.7-fold potetiation when compared with Tc cells obtained from MM-46 tumor-bearing mice which had not received D-fraction (dpm of tumor-bearing D-mice Tc cells-dpm of normal mice Tc cells/dpm of tumor-bearing mice Tc cells-dpm of normal mice Tc cells). However, administration of D-fraction did not potentiate cytotoxic activity against MH-134 cells, having a same major histocompatibility complex(MHC), or against IMC cells, having a different MHC between the tumor cells and host cells. This indicates the activation of Tc cells with antigen-specific cytotoxicity due to the administration of D-fraction. In general, this Tc cell is due to Lyt-2+ T cells, while delayed-type hypersensitivity (DTH) response , on the other hand, is associated with Lyt-1+ T cells.
Consequently, we examined whether the DTH response is actually associated with tumor growth inhibition, and how D-fraction influences DTH response. Lyt-1+ T cells were obtained by removing Lyt-2+ T cells by treatment with anti-Lyt-2.1 monoclonal antibody and complement. A system exclusively containing this T cell fraction or also containing MΦ obtained from normal
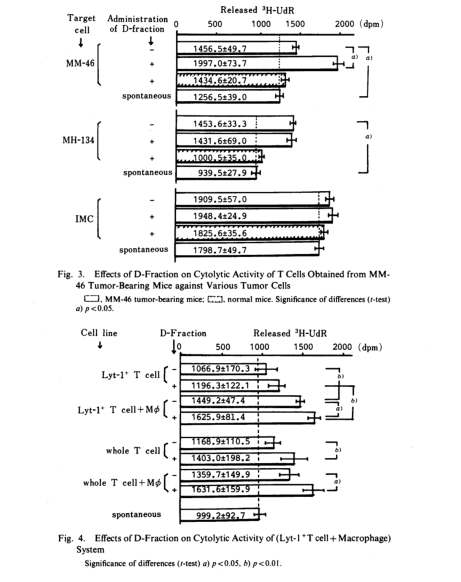
mouse were prepared, and to each was added 3H-uridine-labeled MM-46 tumor cells. After incubation for 48h, released radioactivity was measured. The results are illustrated in Fig.4. Although Lyt-2+ T cells alone exhibited no cytotoxicity in either the D-fraction-administered group of the D-fraction-nonadministered group, cytotoxicity was observed in the presence of normal mouse MΦ which exhibit no cytotoxicity alone. Futhermore, this cytotoxicity was potentiated by the administration of D-fraction. These results suggest that DTH response is associated with tumor growth suppression and that the response is enhanced by D-fraction. When the same experiment was conducted using whole T cells, without separating Lyt-1+ and Lyt-2+ T cells, the potentiation of cytotoxicity was enhanced even in the absence of MΦ. This can be interpreted as resulting from a concented action between Tc cells of Lyt-2+ and DTH-inducing T cells of Lyt-1+ (Tdh cells). These results indicate that the immune systems were stimulated by D-fraction.

Thus, we examined the influence of D-fraction on the production of interleukin-1 (IL-1). The activity of IL-1 in the supernatant of cultured MΦ obtained from D-mice or from tumor-bearing saline-dosed mice was measured in terms of the uptake of 3H-thymidine by T cell obtained from normal mouse. Table V indicates the results. It is seen that 3.2-fold potentiation of IL-1 activity was attained by the administration of D-fraction. This result suggests that the increase in MΦ IL-1 production promoted the activation of the T cell group.
Discussion
Generally, it is well known that some polysaccharides obtained from basidiomycetes spp. show immune-stimulating activity.12) Therefore, the concentrations of the extractable material were expressed in terms of polysaccharide contents. The D-fraction at a dose of 0.75 mg of polysaccharide exhibited antitumor activity. Since the fractionation from pre-A through D-fraction executed in this study followed the procedures adopted in the extraction of alkali-soluble, but acid-insoluble polysaccharide (β-glucan), the fact that D-fraction obtained by this method exhibited a tumor growth-suppressing effect means that a component having an antitumor activity was purified stepwise until an effective concentration was reached at the D-fraction. If a more concentrated solution were administered, other fraction might exhibited a similar effect. However, judging from the concentrations used in this study (all the solutions used for oral administration were viscous) a higher concentration would be difficult to give orally.
On the other hands, we confirmed the presence of a substance which inhibited immune stimulation in the pre-A fraction. It appears that this substance may have been gradually removed in the subsequent fractionations. The D-fraction was further fractionated into E,F and residual fractions. The obtained E-fraction corresponds to MT-2,1) which exhibits a strong antitumor activity when injected intraperitoneally, but proved ineffective when given orally. This dependence on the effect on the route of administration remains to be investigated. Next, the Winn assay was employed to examine whether the tumor-suppressing effect of the D-fraction is due to stimulation of the immune system. As shown in Table VI, when a mixture of tumor cells and spleen cells obtained from a tumor-bearing mouse was injected, significantly higher TIR was obtained as compared with the case of mice inoculated with tumor cells alone. This result suggested that the tumor-bearing mice showed an immune response to the tumor , inducing spleen cells which have cytotoxicity against the tumor cells. Since this action was further potentiated by the administration of D-fraction and a complete inhibition of tumor growth resulted, we assumed that the oral administration of D-fraction strongly activated the cellular immune system which was stimulated by tumor-implantation. Thus, the cytotoxic activity was examined in an in vitro system. When D-fraction was administered, MΦ, T cells and Tdh cells were all activated significantly as compared with the control mouse. As shown in Fig. 3, the activity of Tc cells, which exhibit antigen-specific cytotoxicity, is potentiated, whereas natural killer cells or lymphokine-activated killer cells which exhibit antigen-nonspecific activity, are not influenced. Figure 4 illustrates the results on the cytotoxicity induced by DTH response. Namely, Lyt-1+2- T cells obtained from tumor-bearing mouse showed practically no cytotoxicity alone, but when MΦ, exhibiting no cytotoxicity, were added to this system, cytotoxicity was observed. Futher, in a system consisting of Lyt-1+2- obtained from D-mouse and MΦ, this action was significantly potentiated. These results that Tdh cells, induced in the timor-bearing state, produced MΦ activating factor through contact with antigen tumor cells, and that DTH response was promoted by D-fraction dosing. If such a DTH response could occur also the in vivo system, macrophage activating factor production would be initiated through antigen-specific Tdh response, and the activated MΦ would nonspecifically show cytotoxicity against heterologous cells surrounding the MΦ.
Solid tumors in a host have cell surface antigens.13) Fujiwara et al. ,suggested the possibility that DTH response-activated MΦ play a greater role in antitumor activity than Tc cells. having a higher antigen-specificity14). If so, it would be important to observe the DTH response against tumor cells after the administration of D-fraction. The next phase of the present experiment was to examine interleukin-1 production caused by the administration of D-fraction. As shown in Table V, production of IL-1 was increased by D-fraction, and this IL-1 was considered to act on T cells, promoting the production of IL-2, and elevating the induction of Tc cells or Tdh cells, thus stimulating the whole immune system.
Although the mechanism involved is yet to be elucidated, the fact that D-fraction actually enhanced the production of IL-1 by MΦ collected from tumor-bearing mouse may give a clue to the settlement of this problem. It is believed that gut-associated lymphoid tissue (GALT) is present in the digestive tract. In GALT, T cells, along with B cells, are present.15) Moreover, it was reported that phagocytic epithelial cells in the epithelium of the digestive lumen, together with M cells and MΦ existing in Peyer’s patch, exhibit phagaocytic and antigen-presenting activities.16) Therefore, D-fraction is considered to be subjected to phagocytosis by these cells, resulting in their activation and leading to the promotion of IL-1 production by MΦ existing in GALT, followed by an increase in the production of IL-2. The IL-2 thus produced is considered to enter the blood circulation and then to be associated with the stimulation of the immune system. We intend to investigate the influence of D-fraction on MΦ obtained from the intestinal lumen to obtain evidence for the above suggestion.
Acknowledgement The authors wish to thank Dr. Kanichi Mori and Mr. Tetsuro Toyomasu, Mushroom Research Institute of Japan, for supplying Maitake.
References
1、H. Nanba, A. Hamaguchi and H. Kuroda, Chem. Pharm. Bull., 35,1162 (1987).
2、 K. Adachi, H. Nanba and H. Kuroda, Chem. Pharm. Bull., 35, 261 (1987).
3、N. Ohno, K. Ito, T. Takeyama, I. Suzuki, K. Sato, S. Oikawa and T. Miyazaki, Chem. Pharm. Bull., 33,3395 (1985).
4、T.Mizuno, K.Ohsawa, N.Hagiwara and R.Kuboyama, Agric. Biol. Chem.., 50,1679(1986).
5、H. Nanba, K. Mori, T. Toyomasu and H. Kuroda, Chem. Pharm. Bull., 35, 2453 (1987).
6、H. Nanba and H.Kuroda, Chem. Pharm. Bull., 35, 2459 (1987).
7、T. Ohkuma, T. Otagiri and S. Tanaka, J.Pharmacobio-Dyn.,5, 439 (1982).
8、G. Chihara, J. Hamuro, Y, Arai and F. Fukuoka, Cancer Res., 30, 2776 (1970).
9、S.Fuyama, H.Yamamoto and S.Arai, Cancer Res., 45, 4103 (1985).
10、Y. Izumi, T. Tsuchida, K. Okuno, H. Fujiwara and T. Hamaoka, Gann, 76, 863 (1985).
11、H. Nakajima, Y. Kita, T. Takashi, M. Akasaki, F. Yamaguchi, S. Ozawa, M. Tsukada, S. Abe and D. Mizuno, 75, 260 (1984).
12、G. Chihara, “Gann To Meneki,” Kodansha Scientific, Tokyo, 1981, p. 80.
13、J. L. Urban, M. L. Kripke and H. Schreiber, J.Immunol., 137, 3036 (1986).
14、K. Sakamoto, H. Fujiwara, H. Nakajima, T. Yoshida, Y. Takai and T. Hamaoka, Gann, 77, 1142 (1986).
15、Y. Yoshida, S. Kuroe, M. Chiba, H. Takemori, T. Tsushima, Y. Suwa and A. Kimura, The Saishin-Igaku, 73, 1115 (1982).
16、H. Nakura and K. Tamaki, The Saishin-Igaku, 37, 1053 (1982).
本文部分内容来源网络,如有侵权请联系删除,谢谢。
本文部分内容来源网络,如有侵权请联系删除,谢谢。
编辑:中润世源
